
by Shelby Louis (College of Staten Island, Psychology, 2022-2023 CRSP cohort)
The work was done as a part of the CRSP program at College of Staten Island/CUNY, under the supervision of Dr. Leora Yetnikoff.
This article has been published as part of the Special Edition of Ad Astra, which features the CUNY Research Scholars Program (CRSP) across The City University of New York. The issue is accessible at http://www.adastraletter.com/2024/crsp-special-edition/.
ABOUT THE AUTHOR

Shelby Louis
Shelby Louis is a pre-dental student pursuing a psychology degree at CUNY College of Staten Island. During her junior year she started participating in the CRSP research program and became interested in conducting research and data. Her increased interest opened the door to other researchers and new ideas. Besides her academia, Shelby enjoys her connections with her friends and family enjoys reading a variety of genres in literature.
The corpus callosum is the largest white matter tract of the brain and connects the two hemispheres, ensuring the same information is rapidly and efficiently transmitted to both sides of the brain (Walhovd et al. 2014). Although considered heavily myelinated, hence getting its white appearance, the corpus callosum in both human and rodents in fact only partially myelinated, with some axons unmyelinated and others myelinated inconsistently (Sturrock 1980; Walhovd et al. 2014). Myelin in the brain is produced by a particular glial cell type, the oligodendrocyte (OL), which itself is a differentiated product of self-renewing progenitor cells, oligodendrocyte precursor cells (OPCs). OLs produce layers of lipid which then wrap around axons, providing layers of insulation and enabling the quick transmission of information across long distances with fidelity (Wang et al. 2020). Changes to patterns of axon myelination are believed to affect the synchronization of information coming from distant brain regions (Etxebarria et al. 2016; Noori et al. 2020; Pajevic et al. 2014) and deficits in axon myelination have been proposed to contribute to neuropathology and neuropsychiatric disorders (Ren et al. 2013). Recently, discoveries demonstrated OPCs receive synaptic inputs (Kukley et al. 2007; Ziskin et al. 2007) and that behavioral experiences can affect patterns of myelination by regulating OPC and/or OL activity (Gibson et al. 2014; Marques et al. 2016; Liu et al 2016; Makinodan et al. 2012). However, the neurochemical processes underlying myelin plasticity are not fully known. Because many dopamine-related neurological and neuropsychiatric disorders are also associated with deficits in myelination (Hampton et al. 2019; Karlsgodt 2016; Koshiyama et al. 2018; Lee et al. 2013; Lim et al. 2008; Ma et al. 2009; Mighdoll et al. 2015; Moeller et al. 2005; Rotarska-Jagiela et al. 2008; Samartzis et al. 2014; Tamnes and Agartz 2016; Xu and Li 2011), we hypothesized that dopamine may play a role in myelination plasticity, particularly in the corpus callosum. In support of this hypothesis, we previously demonstrated that functional midbrain dopamine axons are contained within the corpus callosum (Caldwell et al. 2023). We also wanted to investigate whether OPCs and OLs exhibit the mRNA transcripts for dopamine receptors to determine whether these cells contain the necessary machinery to respond to dopamine. To address this, we conducted RNAscope experiments, described here, to determine whether these cell types express mRNA transcripts for dopamine d1(Drd1) and/or dopamine d2(Drd2) receptors. We opted for this method instead of antibody immunofluorescence because there are no reliable antibodies for these receptors.
Perfusions and immunofluorescence
Wild-type (WT) C57/BL6 mice were bred in the Animal Facilities of the College of Staten Island. Mice were maintained on a 12-h light–dark cycle (light on at 0700 h) and given ad libitum access to food and water. Pups were weaned at postnatal day (PND) 21 ± 1 and housed with same-sex littermates. Adult WT (PND 90 ± 15, n = 4; 2 males and 2 females) received an i.p. injection of ketamine/xylazine and were perfused intracardially with 50 ml of 0.9% chilled saline followed by 75 ml of chilled 4% paraformaldehyde in phosphate-buffered saline. Brains were dissected and post-fixed overnight in the same fixative solution at 4°C. Brains were then transferred to phosphate-buffered saline until sectioned using a vibratome.
RNAscope fluorescence in situ hybridization
An in-situ hybridization (ISH) screening for house-keeping genes was performed on coronal 50-µm-thick brain sections from male and female WT mice, pre-mounted onto slides prior to the ISH assay, using RNAScope technology (ACD Bio, Hayward CA, USA). The ISH probes were directed towards Rn-Polr2a (a housekeeping gene), Drd1, Drd2, Olig2 (Oligodendrocyte; marker of OLs), and Pdgfra (Platelet-derived growth factor receptor A; marker of OPCs). Briefly, sections were first fixed in 4% paraformaldehyde for 15 minutes at 4°C, dehydrated in increasing gradients of ethanol baths and left to air dry for 5 minutes. Immunohistochemistry was performed afterward for detection of Olig2, Pdgfra, Drd1, and Drd2 mRNA followed by protease digestion for 30 minutes at 40°C.
Image acquisition and analysis
Immunofluorescence images of the anterior corpus callosum were acquired using a Leica DM6 THUNDER microscope. Regions of interest were then imaged with 20X objectives and the 350-, 488-, 594-, or 647 nm channels. Images were taken in steps of 0.22 to subtend the focal plane of the immunofluorescence signal. Stitched images were then imported into Imaris, a sophisticated semi-automated microscopy image analysis software. A minimum of 2 sections per animal were used. The expression of Drd1 and Drd2 mRNA by all cells within the region of interest expressing Olig2 and/or Pdgfra mRNA was quantified semi-automatically using Object Detection and Nearest Distance features. Specifically, DAPI+, Olig2+, Pdgfra+, Drd1+, and Drd2+ cells. The number of colocalizing structures (defined as zero-0.5 distance in µm) between these different Surface categories was then determined. Sample data that did not satisfy the inclusion criteria were not included in the analysis.
Statistical analysis
RNAScope data are normally distributed and expressed as means ±SEM. Quantitative differences in colocalization of dopamine receptor transcripts by oligodendrocyte lineage cells were analyzed using a 2 (oligodendrocyte lineage cell type) x2 (dopamine receptor subtype) mixed ANOVA.
Upon quantifying the expression of Drd1 and Drd2 mRNA by all Olig2+/Pdgfra+ (OPCs) and Olig2+/Pdgfra- (putative OLs), we discovered that approximately 40% of Olig2+/Pdgfra+ cells expressed Drd1 and Drd2 mRNA transcripts (see also Caldwell et al. 2023). In contrast, only 20% of Olig2+/Pdgfra- cells expressed Drd1 and Drd2 mRNA transcripts, suggesting that the transcript levels are higher in OPCs and decrease as these cells differentiate into OLs (Caldwell et al. 2023). It is important to note that the number of mRNA transcripts expressed by oligodendrocyte lineage cells in the corpus callosum is noticeably less compared to non-oligodendrocyte lineage cells in the nearby medial prefrontal cortex and striatum. However, oligodendrocyte lineage cells in these regions also express Drd1 and Drd2 transcripts at levels similar to those observed in the corpus callosum (Fig. 2).
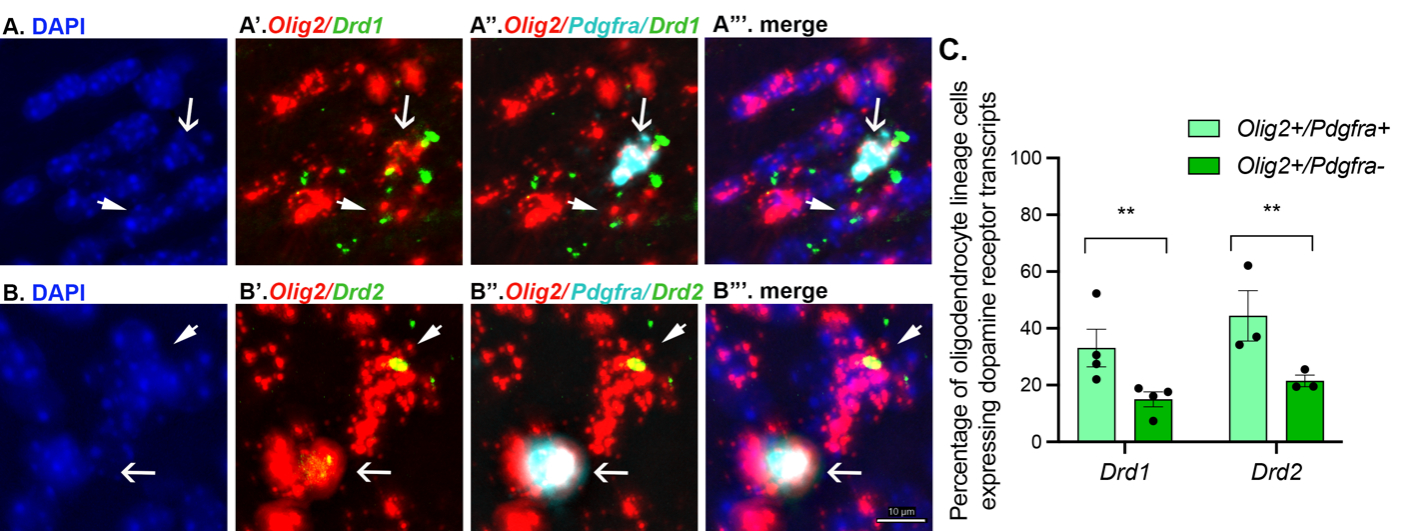
Figure 1: The series of micrographs (A-A’”; B-B’”) demonstrate colocalization of oligodendroglial cells with dopamine receptor transcripts. Olig2+/Pdgfra+ cells reflect OPCs while Olig2+/Pdgfra- cells reflect OLs. Arrows indicate colocalizing cells. The bar graph (C) demonstrates the quantification of the data. In the corpus callosum, there are a greater number of OPCs that express transcripts than OLs. On average, there are nearly twofold more Olig2+Pdgfra+ cells than Olig2+/Pdgfra- expressing dopamine receptor mRNA transcripts. Each black dot represents the average of 2-4 brain section per animal. Figure reproduced from (Caldwell et al. 2023).
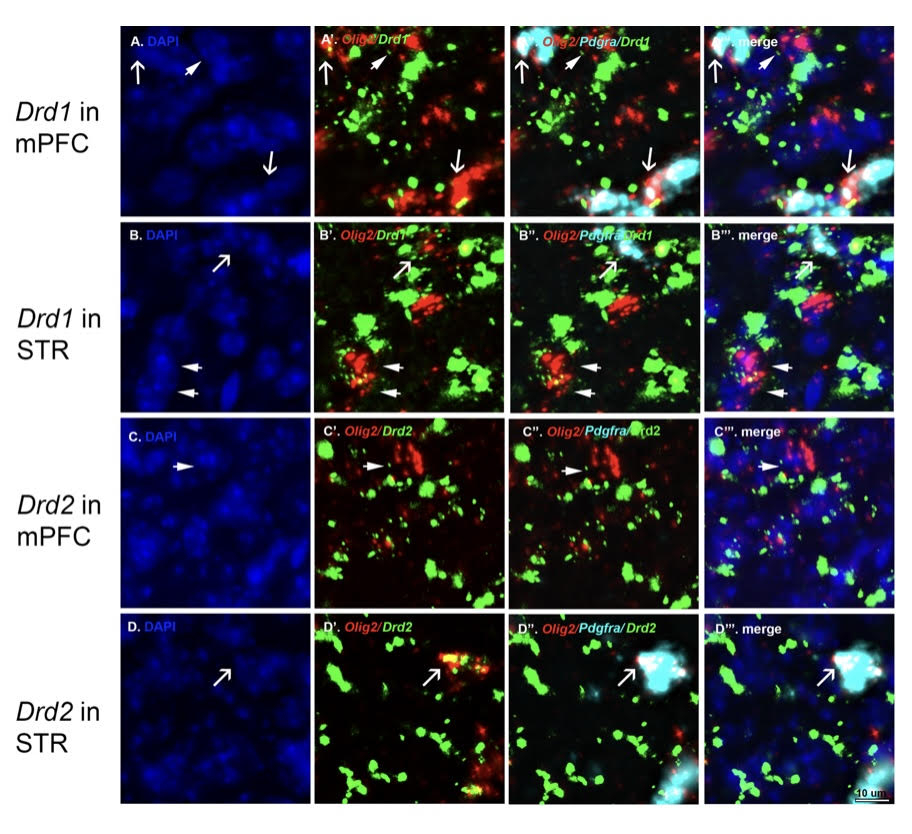
Figure 2: Expression of Drd1 and Drd2 receptor transcripts in the medial prefrontal cortex (mPFC) and striatum. Leica THUNDER images demonstrating DAPI (A - D); Olig2 and Drd1 (A', B') or Olig2 and Drd2 (C', D') RNA probes; Olig2, Pdgfra and Drd1 (A’’, B’’) or Olig2, Pdgfra, Figure 2: Expression of Drd2 (C’’, D’’) RNA probes; and the merge of all channels (A’’’ – D’’’). Note the difference in levels of Drd1 and Drd2 transcript expression compared to the corpus callosum (Figure 1). Intriguingly, some Olig2+/Pdgfra+ cells (arrows) and Olig2+/Pdgfra- cells (arrowheads) in the mPFC and striatum also express Drd1 and Drd2 transcripts. Figure reproduced from (Caldwell et al. 2023).
To our knowledge, we present the first evidence to date supporting the hypothesis that Drd1 and Drd2 receptor mRNA transcripts are expressed in oligodendroglial lineage cells within the corpus callosum. So, not only are functional dopamine axons present within this white matter tract (Caldwell et al. 2023), but we now show that local oligodendrocyte lineage cells, the key players in myelin production, express dopamine receptor transcripts. Although the precise role of dopamine-oligodendroglial communication in the corpus callosum is yet unknown, our studies suggest one role may be in myelin regulation and plasticity, although other non-myelin related roles may also be at play (Caldwell et al. 2023). According to earlier theories, midbrain dopamine neurons are uniquely positioned to play a crucial role in the synchronization of parallel information streams because they are the target of a brain-wide network of afferent inputs and have wide efferent projections (Yetnikoff et al. 2014; Beeler and Dreyer 2019). Their potential contribution to information synchronization could be via the control of myelin plasticity. Dopamine-stimulated changed in intracellular calcium mobilization within oligodendroglial cells may be a potential mechanism in this phenomenon (Undieh 2010). Nevertheless, it will be important to consider other possible roles such as neuron activity-induced changes in cell proliferation, differentiation, and/or even non-myelin related functions such as providing metabolic support or mediating neuro-immune interactions (Xin and Chan 2020). More research is required to understand how oligodendroglial cells express dopamine receptor transcripts in various brain regions and the role played by dopamine receptor signaling in these cells. The implications of this research for comprehending the mechanisms behind brain function and disease, such as multiple sclerosis and schizophrenia, are significant.
This work was conducted under the supervision of Dr. Leora Yetnikoff. Many thanks to the students who equally participated in this research: Josue Criolla Mendoza, Xin Yan Zhou, and Megan Caldwell. KB was funded by Parkinson’s UK (G-1803); LMR was sup- ported the National Institute on Drug Abuse at the National Institute of Health (F31DA041188); SC was funded by the Wellcome Trust (223202/Z/21/Z), Aligning Science Across Parkinson’s (ASAP- 020370), and Medical Research Council (MR/V013599/1); LY was funded by the National Institute of Neurological Disorders and Stroke at the National Institute of Health (1SC2NS125021).
Beeler, J. A., & Kisbye Dreyer, J. (2019). "Synchronicity: The Role of Midbrain Dopamine in Whole-Brain Coordination." Eneuro, 6(2), ENEURO.0345-18.2019. https://doi.org/10.1523/eneuro.0345-18.2019.
Caldwell, M., Ayo-Jibunoh, V., Mendoza, J., Brimblecombe, K. R., Reynolds, L. M., Jiang, X., Alarcón, C., Fiore, E., Tomaio, J. N., Phillips, G. R., Mingote, S., Flores, C., Patrizia Casaccia, Liu, J., Cragg, S. J., McCloskey, D. P., & Yetnikoff, L. (2023). "Axo-glial interactions between midbrain dopamine neurons and oligodendrocyte lineage cells in the anterior corpus callosum." Brain Structure & Function, 228(8), 1993–2006. https://doi.org/10.1007/s00429-023-02695-y.
Daisuke Koshiyama, Fukunaga, M., Okada, N., Morita, K., Nemoto, K., Yamashita, F., Hidenaga Yamamori, Yasuda, Y., Fujimoto, M., Kelly, S., Jahanshad, N., Kudo, N., Hirotsugu Azechi, Watanabe, Y., Donohoe, G., Thompson, P. M., Kasai, K., & Hashimoto, R. (2018). "Role of frontal white matter and corpus callosum on social function in schizophrenia." Schizophrenia Research, 202, 180–187. https://doi.org/10.1016/j.schres.2018.07.009.
Etxeberria, A., Hokanson, K. C., Dao, D. Q., Mayoral, S. R., Mei, F., Redmond, S. A., Ullian, E. M., & Chan, J. R. (2016). "Dynamic Modulation of Myelination in Response to Visual Stimuli Alters Optic Nerve Conduction Velocity." Journal of Neuroscience, 36(26), 6937–6948. https://doi.org/10.1523/JNEUROSCI.0908-16.2016.
Gibson, E. M., Purger, D., Mount, C. W., Goldstein, A. K., Lin, G. L., Wood, L. S., Inema, I., Miller, S. E., Bieri, G., Zuchero, J. B., Barres, B. A., Woo, P. J., Vogel, H., & Monje, M. (2014). "Neuronal Activity Promotes Oligodendrogenesis and Adaptive Myelination in the Mammalian Brain." Science, 344(6183), 1252304–1252304. https://doi.org/10.1126/science.1252304.
Hampton, W. H., Hanik, I. M., & Olson, I. R. (2019). "Substance abuse and white matter: Findings, limitations, and future of diffusion tensor imaging research." Drug and Alcohol Dependence, 197, 288–298. https://doi.org/10.1016/j.drugalcdep.2019.02.005.
Karlsgodt, K. H. (2016). "Diffusion Imaging of White Matter in Schizophrenia: Progress and Future Directions." Biological Psychiatry: Cognitive Neuroscience and Neuroimaging, 1(3), 209–217. https://doi.org/10.1016/j.bpsc.2015.12.001.
Kukley, M., Capetillo-Zarate, E., & Dietrich, D. (2007). "Vesicular glutamate release from axons in white matter." Nature Neuroscience, 10(3), 311–320. https://doi.org/10.1038/nn1850.
Lim, K. O., Wozniak, J. R., Mueller, B. A., Franc, D. T., Specker, S. M., Rodriguez, C. P., Silverman, A. B., & Rotrosen, J. P. (2008). "Brain macrostructural and microstructural abnormalities in cocaine dependence." Drug and Alcohol Dependence, 92(1-3), 164–172. https://doi.org/10.1016/j.drugalcdep.2007.07.019.
Liu, J., Dietz, K., DeLoyht, J. M., Pedre, X., Kelkar, D., Kaur, J., Vialou, V., Lobo, M. K., Dietz, D. M., Nestler, E. J., Dupree, J., & Casaccia, P. (2012). "Impaired adult myelination in the prefrontal cortex of socially isolated mice." Nature Neuroscience, 15(12), 1621–1623. https://doi.org/10.1038/nn.3263.
Ma, L., Hasan, K. M., Steinberg, J. L., Narayana, P. A., Lane, S. D., Zuniga, E. A., Kramer, L. A., & Moeller, F. G. (2009). "Diffusion tensor imaging in cocaine dependence: Regional effects of cocaine on corpus callosum and effect of cocaine administration route." Drug and Alcohol Dependence, 104(3), 262–267. https://doi.org/10.1016/j.drugalcdep.2009.05.020.
Makinodan, M., Rosen, K. M., Ito, S., & Corfas, G. (2012). "A Critical Period for Social Experience–Dependent Oligodendrocyte Maturation and Myelination." Science (New York, N.Y.), 337(6100), 1357–1360. https://doi.org/10.1126/science.1220845.
Marques, S., Zeisel, A., Codeluppi, S., van Bruggen, D., Mendanha Falcao, A., Xiao, L., Li, H., Haring, M., Hochgerner, H., Romanov, R. A., Gyllborg, D., Munoz-Manchado, A. B., La Manno, G., Lonnerberg, P., Floriddia, E. M., Rezayee, F., Ernfors, P., Arenas, E., Hjerling-Leffler, J., & Harkany, T. (2016). "Oligodendrocyte heterogeneity in the mouse juvenile and adult central nervous system." Science, 352(6291), 1326–1329. https://doi.org/10.1126/science.aaf6463.
McKenzie, I. A., Ohayon, D., Li, H., Paes de Faria, J., Emery, B., Tohyama, K., & Richardson, W. D. (2014). "Motor skill learning requires active central myelination." Science, 346(6207), 318–322. https://doi.org/10.1126/science.1254960.
Mighdoll, M. I., Tao, R., Kleinman, J. E., & Hyde, T. M. (2015). "Myelin, myelin-related disorders, and psychosis." Schizophrenia Research, 161(1), 85–93. https://doi.org/10.1016/j.schres.2014.09.040.
Moeller, F. G., Hasan, K. M., Steinberg, J. L., Kramer, L. A., Dougherty, D. M., Santos, R. M., Valdes, I., Swann, A. C., Barratt, E. S., & Narayana, P. A. (2004). "Reduced Anterior Corpus Callosum White Matter Integrity is Related to Increased Impulsivity and Reduced Discriminability in Cocaine-Dependent Subjects: Diffusion Tensor Imaging." Neuropsychopharmacology, 30(3), 610–617. https://doi.org/10.1038/sj.npp.1300617.
Noori, R., Park, D., Griffiths, J. D., Bells, S., Frankland, P. W., Mabbott, D., & Lefebvre, J. (2020). "Activity-dependent myelination: A glial mechanism of oscillatory self-organization in large-scale brain networks." Proceedings of the National Academy of Sciences, 117(24), 13227–13237. https://doi.org/10.1073/pnas.1916646117.
Pajevic, S., Basser, P. J., & Fields, R. D. (2014). "Role of myelin plasticity in oscillations and synchrony of neuronal activity." Neuroscience, 276, 135–147. https://doi.org/10.1016/j.neuroscience.2013.11.007.
Rotarska-Jagiela, A., Ralf Schönmeyer, Oertel, V., Haenschel, C., Vogeley, K., & Edmund, D. (2008). "The corpus callosum in schizophrenia-volume and connectivity changes affect specific regions." NeuroImage, 39(4), 1522–1532. https://doi.org/10.1016/j.neuroimage.2007.10.063.
Samartzis, L., Dima, D., Fusar-Poli, P., & Kyriakopoulos, M. (2013). "White Matter Alterations in Early Stages of Schizophrenia: A Systematic Review of Diffusion Tensor Imaging Studies." Journal of Neuroimaging, 24(2), 101–110. https://doi.org/10.1111/j.1552-6569.2012.00779.x.
STURROCK, R. R. (1980). "MYELINATION OF THE MOUSE CORPUS CALLOSUM." Neuropathology and Applied Neurobiology, 6(6), 415–420. https://doi.org/10.1111/j.1365-2990.1980.tb00219.x.
Tamnes, C. K., & Agartz, I. (2016). "White Matter Microstructure in Early-Onset Schizophrenia: A Systematic Review of Diffusion Tensor Imaging Studies." Journal of the American Academy of Child & Adolescent Psychiatry, 55(4), 269–279. https://doi.org/10.1016/j.jaac.2016.01.004.
Undieh, A. S. (2010). "Pharmacology of signaling induced by dopamine D1-like receptor activation." Pharmacology & Therapeutics, 128(1), 37–60. https://doi.org/10.1016/j.pharmthera.2010.05.003.
Walhovd, K. B., Johansen-Berg, H., & Káradóttir, R. T. (2014). "Unraveling the secrets of white matter – Bridging the gap between cellular, animal and human imaging studies." Neuroscience, 276, 2–13. https://doi.org/10.1016/j.neuroscience.2014.06.058.
Wang, J., He, X., Meng, H., Li, Y., Dmitriev, P., Tian, F., Page, J. C., Lu, Q. R., & He, Z. (2020). "Robust Myelination of Regenerated Axons Induced by Combined Manipulations of GPR17 and Microglia." Neuron, 108(5), 876-886.e4. https://doi.org/10.1016/j.neuron.2020.09.016.
Xin, W., & Chan, J. R. (2020). "Myelin plasticity: sculpting circuits in learning and memory." Nature Reviews Neuroscience, 21(12), 682–694. https://doi.org/10.1038/s41583-020-00379-8.
Yetnikoff, L., Lavezzi, H. N., Reichard, R. A., & Zahm, D. S. (2014). "An update on the connections of the ventral mesencephalic dopaminergic complex." Neuroscience, 282, 23–48. https://doi.org/10.1016/j.neuroscience.2014.04.010.
Ziskin, J. L., Nishiyama, A., Rubio, M., Fukaya, M., & Bergles, D. E. (2007). "Vesicular release of glutamate from unmyelinated axons in white matter." Nature Neuroscience, 10(3), 321–330. https://doi.org/10.1038/nn1854.