by Michael Betancourt and Dr. Charles Keller
ABOUT THE AUTHORS

Dr. Charles Keller
Charles Keller is a Professor of Biology in the Natural Sciences Department at LaGuardia Community College. He began teaching at LaGuardia in 2012. He received his Ph.D. from Columbia University in New York in 2011 where he studied the development and function of Touch Receptor Neurons (TRNs) in the soil-dwelling nematode Caenorhabiditis elegans with an emphasis on the role of microtubules and the regulation of TRN gene expression and terminal differential by the LIM-Homeodomain transcription factor MEC-3. His current research is focused on tardigrades though other projects conducted at LaGuardia include the effect of Bis-phenol A exposure on TRN development in C. elegans and the role of Microtubule Associated Proteins (MAPs) in the C. elegans light touch avoidance response.

Michael Betancourt
Michael Betancourt is currently an upper senior at CUNY Queens College and is an incoming neuroscience Ph.D. student at Weill Cornell Graduate School of Medical Sciences. He obtained his associates degree in biology at LaGuardia Community College in 2019 and is soon to finish his bachelor’s degree in psychology at Queens College. During his time at LaGuardia Community College, Michael worked in the laboratory of Dr. Charles Keller studying genetic adaptations to heavy metal toxicity in tardigrades inhabiting highly polluted environments. While attending Queens College, Michael worked in the laboratory of Dr. Maral Tajerian, where he studied hippocampal extracellular matrix plasticity in mice models of chronic pain, and how this plasticity modulates glial-neuronal interactions following injury. Going into graduate school, Michael’s research interests involve studying the extracellular matrix, glial-neuronal interactions, and synaptic plasticity in different models of neurological disorders.
Background
Most known species on Earth survive within a relatively narrow range of environmental parameters. Extremes of temperature, pressure, PH, salinity, radiation, desiccation, and a variety of other variables prove prohibitive for the long-term survival of most species. The concept of a planetary “Goldilocks Zone” summarizes these observations with a simple analogy to the story of Goldilocks and the Three bears. Goldilocks, an interloper in the Three Bear's home, finds that she prefers to pilfer the porridge that is neither too hot nor too cold but just right. Earth is “just right” for life as we understand it to have arisen and persisted: Closer to the sun is too hot and further away is too cold for rocky planets, with strong magnetic fields, capable of sustaining liquid water, and thus life, to form.
While scattered examples of particularly hardy species have been known for some time (e.g., Deinococcus radiodurans; Brooks & Murray 1981), advances in submersibles, aerospace, molecular biology, and other technologies in the past several decades have led to the discovery and characterization of a wide variety of species capable of surviving comfortably in environments previously believed to be non-permissive. These particularly hardy species include those found at the bottom of the ocean under crushing pressures and in abysmal darkness, deep in caves far from the sun, buried under kilometers of Antarctic ice, and in near boiling water. These hardy organisms are of particular interest to scientists studying the possibility of extraterrestrial life as they expand the list of reasonable places to look. Ongoing discovery and characterization of these hardy species has forced a reconsideration of the boundaries assigned to the Goldilocks Zone and led to the exciting realization that many environments within the solar system may be less hostile to life than was previously believed. The realization that life on our own planet is able to occupy almost any available niche coupled with the deluge of newly discovered, extra-solar planets in the past two decades makes this a particularly exciting time for the field of Astrobiology. An understanding of the evolution and molecular basis of particularly hardy species may provide insight into the nature of putative extraterrestrial life as well as the rise and diversification of life on primordial Earth.
Members of the phylum Tardigrada are of particular interest to the Astrobiology community due to some species' ability to withstand a variety of extreme terrestrial and low-orbit environments which may be analogous to extraterrestrial or early Earth habitats. Tardigrades (commonly known as “water bears” due to their lumbering, bear-like appearance) are microscopic, aquatic, invertebrates including over 1100 documented species (Bartels et al., 2016). Some Tardigrades can withstand extremes of cold (Hengherr et al., 2009), radiation (Beltrán-Pardo et al., 2015), pressure (Ono et al., 2016), osmolarity (Bjørn-Mortensen, 2006), and desiccation even when in an active metabolic state (Møbjerg et al., 2011). Some Tardigrade species are also capable of anhydrobiosis, a metabolically inactive, desiccated state entered in response to extreme dehydration (Czernekowa & Jönsson et al., 2016). Tardigrades were the first animal to have been shown to survive exposure to the extreme conditions encountered in low Earth orbit (Jönsson et al., 2008) leading to the tantalizing possibility of tardigrade participation in panspermic colonization events as suggested by Georgiev (Georgiev, 2016) and supported by the observation that Hypsibius Dujardini can survive hypervelocity impact events analogous to those experienced upon foreign body impact with planetary surfaces despite being one of the less extremotolerant tardigrades (Pasini, 2014). These features have led to tardigrades being suggested as a possible “model animal” for astrobiological study (Horikawa, 2012).
Pollution caused by human activity may represent relatively sudden evolutionary challenges to organisms in heavily polluted environments. The study of particularly hardy species in heavily polluted environments may lead to the identification of novel adaptations at the genetic level that allow survival in the sort of extreme or fluctuating environments found on early earth or on Extra-terrestrial bodies. A variety of studies have been performed addressing tardigrade responses to polluted environments and urban environments particularly regarding their density and distribution. The bryophytes which represent a common tardigrade habitat and food source are effective bio magnifiers of airborne heavy metals and other pollutants (Markert, 1993) with apparent consequences for tardigrade diversity as species number on bryophyte,lichens, and other habitats has been observed to be inversely correlated with air pollution levels (Vargha et al., 2002, Hohl et al., 2001, Ðatkauskienë, 2012). While the above cited studies and others suggest that exposure to pollutants such as SO2 and heavy metals can result in decreased tardigrade density and species diversity, little is known about tardigrade gene expression changes (or evolutionary adaptation) in response to heterogenous pollutants in an urban context or heavy metal exposure in general. Indeed, a tardigrade copper-tolerance gene expression study performed by Hygum and coworkers in 2017 claims to be the first to investigate heavy metal tolerance in tardigrades (Hygum et al., 2017) We hypothesized that Tardigrades living in or adjacent to Newtown Creek, with its nearly two century history of heavy industrial pollution (Hurley, 1994), might exhibit novel adaptations in genes encoding or controlling the activity of enzymes that protect cells against a variety of pollutants including heavy metals.
We proposed to isolate DNA from Tardigrades living in the Newtown Creek environment and obtain DNA sequences of genes known to be involved in resistance to heavy metal toxicity including members of the Super Oxide Dismustase (SOD) family of enzymes. SODs are an evolutionarily conserved family of metalloenzymes involved in protecting cells from oxidative damage. Heavy metals can outcompete the normal cofactors of SODs (copper, zinc, manganese, etc) and inhibit their activity, thus leading to cellular damage. We speculated that DNA sequences of SODs obtained from Newtown Creek Tardigrades might reveal adaptions to environments containing high levels of heavy metals.
Species identification of tardigrades involves painstaking analysis of a variety of microscopic morphological features and is notoriously challenging. We therefore proposed to identify isolated animals using established molecular phylogeny approaches in addition to visual examination. 18s Ribosomal RNA and Cytochrome Oxidase Subunit I (COI) sequences are found in all eukaryotes but subtle variations in sequence are typically species-specific and can serve as the basis for species identification (Cesari et al., 2013).
Experimental methods
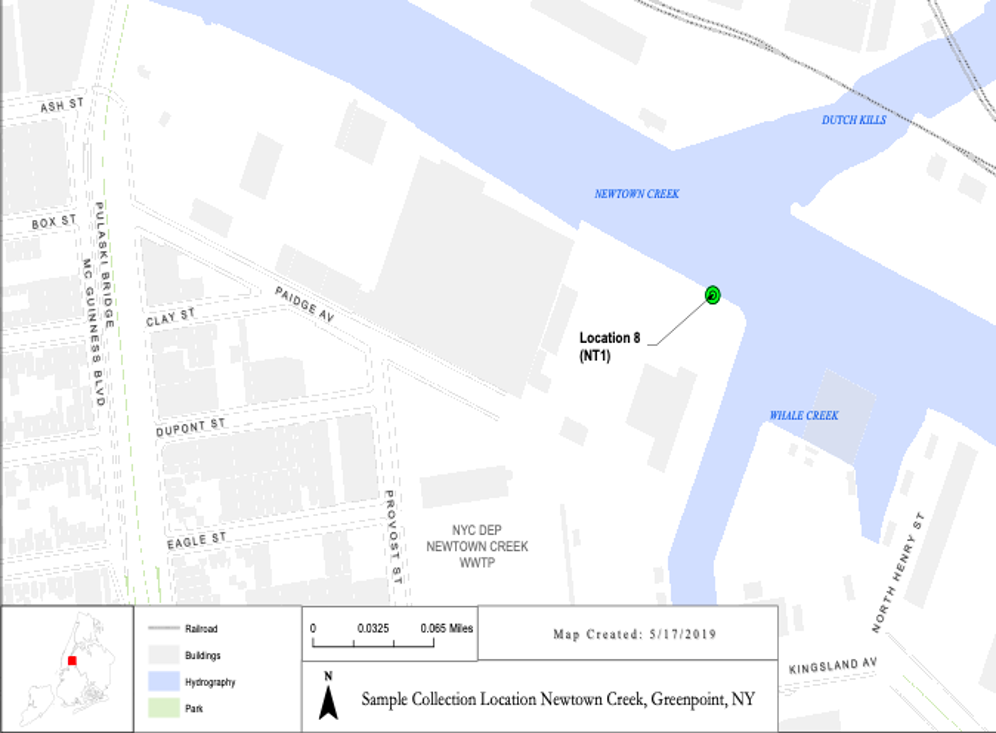
Tardigrade isolation: Samples were collected from moss, barnacles, and cement surfaces above and below the high tide line at the Newtown Creek Nature Walk (Fig. 1) and from lichen or moss growing on trees outside of the M-building. Samples were returned to the laboratory for further examination. Dry samples were hydrated using spring water while water/barnacle shell samples were maintained in ambient water until examination. Samples were transferred to petri dishes and examined using a dissecting microscope. Individual tardigrades were identified and removed (using a micropipette) and placed in separate plates containing a 1% agarose base for ease of movement, spring water, as well as algae (Carolina Biological) as a source of food. 5-10 individuals were moved to each plate in the hopes of establishing a reproducing lab colony. Many tardigrade species are quite similar in appearance and we could not determine if isolated individuals were from the same or different species by visual inspection alone. Single tardigrades were therefore isolated and single-animal DNA extractions and subsequent rRNA or COI PCR were performed to confirm the species composition of any laboratory colonies established.
Single animal DNA extraction and PCR
Single tardigrades were isolated from plates and placed in Eppendorf tubes containing lysis buffer, and a homogenizer was used to break down the sample for DNA extraction (Protocol used: Zymo Research solid tissue sample DNA extraction kit). Extracted DNA was quantified using NANODROP 8000 spectrophotometer and used as a template for Polymerase Chain Reaction (PCR) to amplify cytochrome oxidase I (COI) and 18S rRNA, sequences for species identification (discussed in Cesari et al., 2013). DNA extracted from single animals also served as the template for our SOD PCR reactions. Amplified and gel-confirmed PCR products were sent to Genewiz for PCR purification and Sanger DNA sequencing. Results were compared to known sequences present in the National Center for Biotechnology database using the Blastn sequence alignment tool.
Microscopy
Individual animals were placed on 1% agarose pads on glass slides and covered with a cover slip for microscopy and photography using a Nikon inverted fluorescence Ti-E microscope. Imaged individuals were subsequently moved to eppendorf tubes for single-tardigrade DNA extraction.
Results:
Establishment of lab culture:
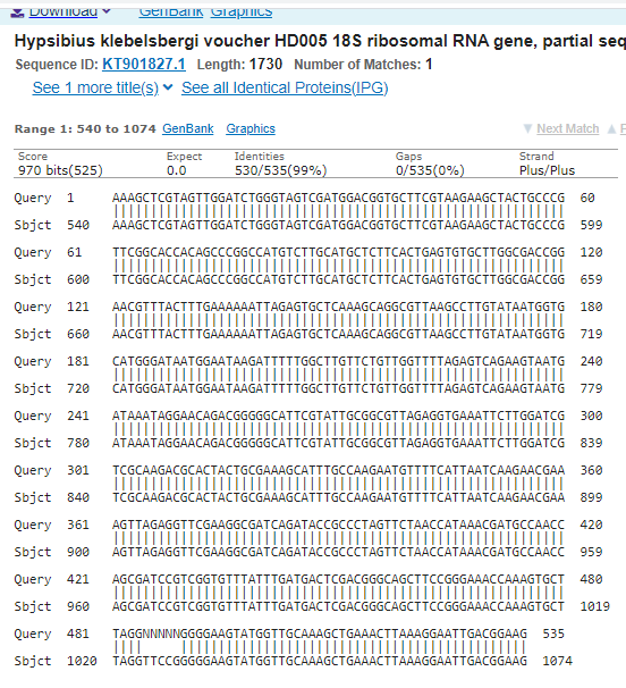
While most of our attempted lab cultures did not survive and reproduce under our conditions, one reproducing lab culture was established from a moss sample taken from the Newtown Creek Nature walk steps. This culture was maintained for over one year at room temperature in Spring water supplemented with an algal food source under constant light (Its current status is unknown due to the pandemic). 18S rRNA sequences obtained from individuals in this culture show an identical match with curated sequences for the tardigrade species Hypsibius klebelsbergi (NCBI:txid2792769 originally Identified as Cryobiotus klebelsbergi (Mihelčič, 1959)) (Fig. 2). While we are hopeful that our colony solely consists of H klebelsbergi, we cannot rule out the possibility of other tardigrade species being present until we are able to obtain more individual tardigrade DNA sequences.
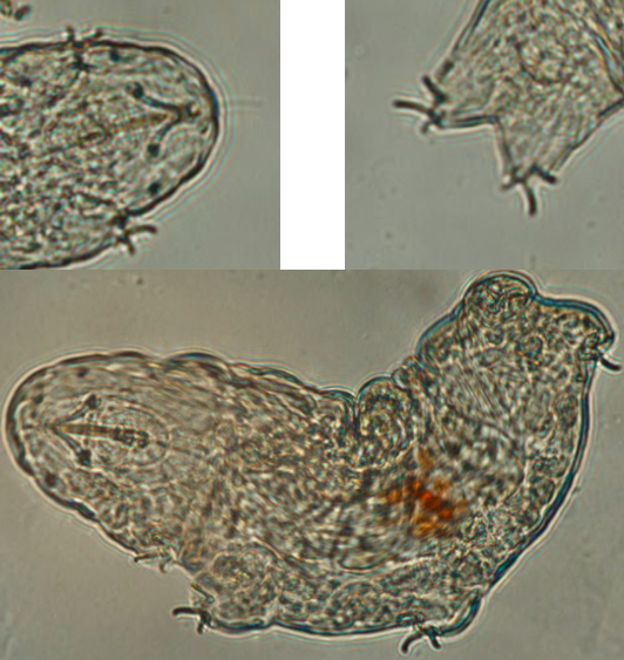
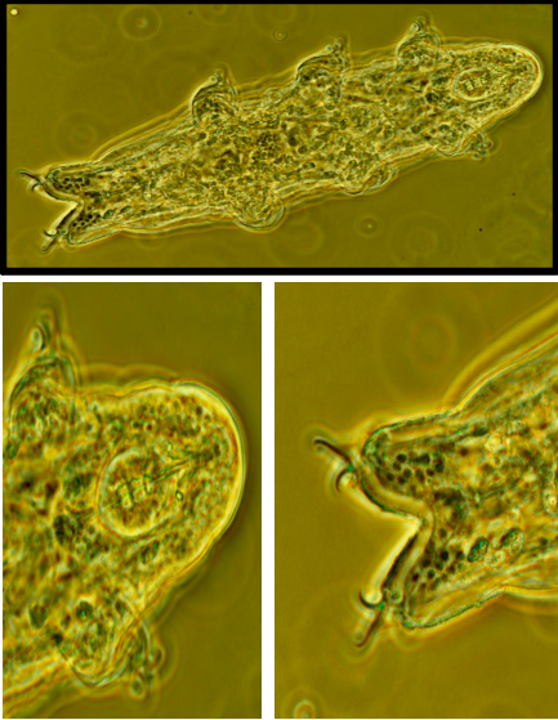
In addition to the sequences mentioned above, individuals of the following species taken from other samples were identified by one DNA sequence match but not successfully cultured Ramazzottius oberhaeuseri (18S ribosomal RNA match. Fig. 3) and a putatively unknown species of similarity to Macrobiotus macrocalux based on COI sequence (Fig. 4).
PCR amplification of known stress-resistance genes:
Degenerate PCR primers were designed based on known Copper/Zinc Super-Oxide Dismutase sequences (The SODs are highly conserved across phyla but no Tardigrade SODs sequences are available in the NCBI database). These primers have thus far failed to produce a PCR product and new primers are being designed.
Microscopy:
We successfully isolated and observed multiple living tardigrades though we have thus far been unable to successfully extract DNA from imaged individuals (which would be ideal for species identification allowing both molecular and morphological data to be combined). We are unable to confidently identify imaged individuals at this time. Representative images from two individuals are shown (Figs 5 and 6).
Conclusions:
We successfully established a laboratory line of tardigrades isolated from the Newtown Creek Superfund shoreline which will allow us to perform a variety of stress resistance assays. We believe this culture to consist exclusively of H. klebelsbergi though further confirmation is needed as this identification is based on DNA sequences from a handful of individual animals. We will perform multiple rounds of replica plating coupled with single-tardigrade PCR to confirm the species composition of our colony. Confirmation of H. klebelsbergi in Newtown Creek would be of particular interest as it has only been previously described in harsh glacial habitats (Dastych, et al., 2003) and has indeed been referred to as “an obligate glacial dweller” (Dabert et al., 2015). We were also able to successfully extract DNA from individual tardigrades and use established molecular phylogeny techniques to strongly suggest species identity. Attempts to image individual tardigrades were successful though we have yet to image individuals and subsequently successfully extract DNA, perform a successful PCR reaction, and obtain a usable sequence from that same animal.
We feel that the results presented above represent exciting preliminary data and proof of principle/technique as the basis for future work. We are particularly excited about the possibility of characterizing a species previously thought to be adapted for harsh glacial environments should our preliminary identification of H klebelsbergi be confirmed. However, we must acknowledge that comprehensive, global surveys of micro-animal diversity are quite difficult, and it could be the case that H klebelsbergi is more widespread than previously known. We look forward to returning to our laboratory and characterizing more of Newtown Creek’s long-suffering tardigrades. Future projects include confirmation of our H. klebelsbergi culture, tardigrade survival assays, gene expression characterization of cultured tardigrades in response to stress, and more sequencing of DNA from individual animals to estimate evolution rates at the level of individual genes.
Acknowledgements: We would like to thank Alyson Sheffield and Joshua Tan from the Natural Sciences Department who brought Tardigrade biology and astrobiology to LaGuardia and provided the inspiration for this work. We would also like to thank the many scientists in the Natural Science department and beyond who have facilitated this work or who have long-running projects involving Newtown Creek including but not limited to: Holly Porter-Morgan, Sarah Durand, Olga Calderon, Joby Jacob, Ingrid Veras, Willis Elkins, The Newtown Creek Alliance, and others. We would also like to thank Irina V. Dopson of the NYC Department of Environmental Protection for assisting with Figure 1. Michael Betancourt was supported by the NIH Bridges Program.
References:
Bartels, P. J., Apodaca, J. J., Mora, C., & Nelson, D. R. (2016). A global biodiversity estimate of a poorly known taxon: phylum Tardigrada. Zoological Journal of the Linnean Society, 178(4), 730-736.
Beltrán-Pardo, E., Jönsson, K. I., Harms-Ringdahl, M., Haghdoost, S., & Wojcik, A. (2015). Tolerance to gamma radiation in the tardigrade Hypsibius dujardini from embryo to adult correlate inversely with cellular proliferation. PLoS One, 10(7), e0133658.
Bjørn-Mortensen, M. (2006). Osmoregulation and osmobiosis in the eutardigrade Richtersius coronifer. Roskilde Roskilde University Institute for Biology and Chemistry.
Brooks, B. W., & Murray, R. G. E. (1981). Nomenclature for “Micrococcus radiodurans” and other radiation-resistant cocci: Deinococcaceae fam. nov. and Deinococcus gen. nov., including five species. International Journal of Systematic and Evolutionary Microbiology, 31(3), 353-360.
Cesari, M., Guidetti, R., Rebecchi, L., Giovannini, I., & Bertolani, R. (2013). A DNA barcoding approach in the study of tardigrades.
Czernekova, M., & Jönsson, K. I. (2016). Experimentally induced repeated anhydrobiosis in the eutardigrade Richtersius coronifer. PLoS One, 11(11), e0164062.
Dabert, Miroslawa., Dastych, Hieronymus., & Dabert, Jacek (2015). Molecular data support the dispersal ability of the glacier tardigrade Hypsibius klebelsbergi Mihelčič, 1959 across the environmental barrier (Tardigrada). Entomologische Mitteilungen aus dem Zoologischen Museum Hamburg, 17(194), 233-240.
Ðatkauskienë, I. (2012). Microfauna of lichen (Xanthoria parietina) in Lithuania: diversity patterns in polluted and non-polluted sites. Baltic Forestry, 18, 255-262.
Dastych, Hieronymus., Kraus, H. J., & Thaler, K. (2003). Redescription and notes on the biology of the glacier tardigrade Hypsibius klebelsbergi Mihelcic, 1959 (Tardigrada), based on material from Ötztal Alps, Austria. Mitt. hamb. zool. Mus. Inst, 100, 77-100.
Georgiev, D. (2016). The Phylum Tardigrada and the Panspermia Theory? Can The Tardigrades be Live Capsules Carrying a Variety of DNA Sequences Inside as Food Particles, Endosymbiotic Organisms and Parasites? International Journal of Pure and Applied Zoology, 4(4).
Hengherr S, Brümmer F, Schill R.O. (2008). Anhydrobiosis in tardigrades and its effects on longevity traits. Journal of Zoology, 275:216–220.
Hohl, A. M., Miller, W. R., & Nelson, D. R. (2001). The Distribution of Tardigrades Upwind and Downwind of a Missouri Coal-Burning Power Plant. Zoologischer Anzeiger-A Journal of Comparative Zoology, 240(3-4), 395-401.
Horikawa D.D. (2012). Survival of Tardigrades in Extreme Environments: A Model Animal for Astrobiology. In: Altenbach A., Bernhard J., Seckbach J. (eds) Anoxia. Cellular Origin, Life in Extreme Habitats and Astrobiology, vol 21. Springer, Dordrecht.
Hurley, A. (1994). Creating Ecological Wastelands: Oil Pollution in New York City, 1870-1900. Journal of Urban History, 20(3), 340-364.
Hygum, T. L., Fobian, D., Kamilari, M., Jørgensen, A., Schiøtt, M., Grosell, M., & Møbjerg, N. (2017). Comparative investigation of copper tolerance and identification of putative tolerance related genes in tardigrades. Frontiers in Physiology, 8, 95.
Jönsson, K. I., Rabbow, E., Schill, R. O., Harms-Ringdahl, M., & Rettberg, P. (2008). Tardigrades survive exposure to space in low Earth orbit. Current biology, 18(17), R729-R731.
Markert, B. (1993). Plant as biomonitors-Indicators for heavy metals in the terrestrial environment (Vol. 298). Weinheim, Germany.: VCH Verlagsgesellschaft mbH.
Møbjerg, N., Halberg, K. A., Jørgensen, A., Persson, D., Bjørn, M., Ramløv, H., & Kristensen, R. M. (2011). Survival in extreme environments-on the current knowledge of adaptations in tardigrades. Acta physiologica, 202(3), 409-420.
Ono, F., Mori, Y., Takarabe, K., Fujii, A., Saigusa, M., Matsushima, Y., ... & Saini, N. L. (2016). Effect of ultra-high pressure on small animals, tardigrades and Artemia. Cogent Physics, 3(1), 1167575.
Pasini, D. (2014, April). Survival of the Tardigrade Hypsibius Dujardini during Hypervelocity Impact Events up to 5.49 km s-1. In, European Planetary Science Congress (Vol. 9).
Vargha, B., Ötvös, E., & Tuba, Z. (2002). Investigations on ecological effects of heavy metal pollution in Hungary by moss-dwelling water bears (Tardigrada), as bioindicators. Annals of agricultural and environmental medicine, 9(2), 141-146.